食源性致病菌生長延滯期的影響因素
生長延滯期的影響因素
生長環(huán)境
預測微生物學研究表明,食源性致病菌所處的實際生長環(huán)境會影響生長延滯期的長短。Buchanan等早期的研究表明,單核細胞增生李斯特菌的群體生長延滯期隨實際環(huán)境的溫度和pH值的降低而相應延長,隨后的研究結果也表明E.coliO157:H7的群體生長延滯期隨實際環(huán)境溫度降低而有延長的趨勢。
類似的結果也出現(xiàn)在食源性致病菌單細胞水平的研究中,Parra-Flores等對不同生長溫度下的阪崎克羅諾桿菌單細胞生長延滯期分布進行研究,發(fā)現(xiàn)隨溫度降低,單細胞生長延滯期顯著延長,同時在不利的生長環(huán)境下其變異性更強,從而導致生長延滯期的分布模型曲線范圍更廣。Aguirre等研究不同水分活度(0.940~0.997)下單核細胞增生李斯特菌單細胞生長延滯期的變化情況,結果表明隨水分活度降低單細胞生長延滯期有延長的趨勢。此外,圍繞食品基質中食源性致病菌的生長動力學也開展了較多研究。Sant'ana等研究即食生菜中單核細胞增生李斯特菌及腸炎沙門氏菌在7~30℃范圍內的生長參數(shù)變化情況,并準確建立生長延滯期與貯存溫度關系的二級模型,所采用的平方根模型擬合度良好,能夠很好地描述即食生菜貯存過程中兩種致病菌的動力學行為。同樣地,胡錚瑢和孟云等分別對不同溫度下(6、15、25、35℃)清蛋糕中及不同溫度下(5、10、15、20、25℃)涼皮中金黃色葡萄球菌生長延滯期二級模型進行了構建。綜上,不利的實際生長條件可延長生長延滯期。在對食源性致病菌生長延滯期的研究中,溫度、pH值、水分活度等為主要研究因素,由于在食品生產鏈中溫度波動最大,且對細菌的生長影響作用最顯著,所以溫度是研究最多的因素。不同基質中溫度影響食源性致病菌生長延滯期的相關研究見下表。

不同基質中溫度影響食源性致病菌生長延滯期的相關研究
在微生物生長過程中,生長延滯期是環(huán)境變化引起的生長延遲反應的時期。微生物接種于某一環(huán)境,生長一段時間后轉移至另一環(huán)境繼續(xù)生長。在該過程中,微生物最初的生長環(huán)境為前接種環(huán)境或歷史生長環(huán)境,之后的生長環(huán)境為實際生長環(huán)境。除實際生長環(huán)境外,歷史生長條件對食源性致病菌生長延滯期也存在顯著影響。
在早期的研究中,Dufrenne等研究在歷史生長溫度為7℃和37℃下分別培養(yǎng)35 d和5 d(每周或每天更換新鮮培養(yǎng)液)的蠟樣芽孢桿菌在實際溫度為7℃下的生長延滯期,結果表明歷史生長溫度為7℃的蠟樣芽孢桿菌生長延滯期更短。同樣地,Dykes等對歷史生長溫度為4、20、37℃的單核細胞增生李斯特菌在實際溫度4℃下的生長情況進行比較,結果表明歷史生長溫度為4℃的單核細胞增生李斯特菌較快地進入對數(shù)生長期,表現(xiàn)出更短的生長延滯期。
類似的研究結果出現(xiàn)在Yue Siyuan等采用活菌計數(shù)法觀測不同歷史溫度對單核細胞增生李斯特菌于25℃下生長情況的研究中,隨著歷史溫度與生長溫度之間溫差的降低(25℃降低至10℃),生長延滯期從4.14 h縮短至0.30 h。除單一地研究溫度外,F(xiàn)rancois等探究了歷史生長溫度和pH值對7℃下單核細胞增生李斯特菌單細胞生長的影響,發(fā)現(xiàn)相同歷史生長溫度下,歷史生長pH值從7.4降至5.7,單細胞生長延滯期縮短。Tiganitas等研究證明歷史生長pH值(5.0、7.2)及歷史生長水分活度(0.930、0.995)對單核細胞增生李斯特菌在不同水分活度下的生長延滯期均呈現(xiàn)顯著影響,其中歷史生長水分活度與實際生長水分活度越接近,則生長延滯期越短。綜上,食源性致病菌的生長延滯期不僅受實際及歷史生長條件的影響,還受歷史與實際生長條件間的變化量影響,且變化量減小可縮短生長延滯期。根據(jù)實際中食品加工條件的復雜性,應設計更多的歷史及實際生長條件并量化以上兩種條件對生長延滯期的影響,從而提高生長預測模型的準確性,使其能夠更好地應用于食源性致病菌風險評估中。
亞致死損傷
在食品加工等環(huán)節(jié)中,通過多種加工方法處理后,致病菌可能會出現(xiàn)3種不同生理狀態(tài)的細胞:正常細胞、亞致死損傷狀態(tài)的細胞和死亡細胞。其中亞致死損傷菌不能通過普通檢測致病菌的方法檢測出,而一旦處于適宜的環(huán)境條件下,亞致死損傷菌能夠自我修復并恢復到正常狀態(tài),這將給食品安全帶來一定的風險。由于亞致死損傷菌存在修復過程,生長延滯期要比正常細胞的生長延滯期更長,而生長延滯期的長短與細胞的損傷程度、壓力處理方式、細菌種類、食品的成分及貯存條件均有關。
在眾多食品處理方式中,熱力殺菌是一種應用最早、使用最廣泛且效果可靠的方法,這其中溫和熱加工方法由于具有能夠很好地保持食品營養(yǎng)成分及感官特性等優(yōu)勢已成為熱加工發(fā)展趨勢和研究熱點之一,然而此方法很可能由于加工不充分產生大量處于亞致死損傷狀態(tài)的致病菌。如Xuan Xiaoting等對55℃熱處理后Listeria monocytogenesATCC 19114-3的生長延滯期進行了研究。結果發(fā)現(xiàn)55℃處理10 min后Listeria monocytogenesATCC 19114-3的亞致死損傷率為60.19%,且在15℃修復條件下的生長延滯期與未處理組相比延長了4.74 h。Aguirre等研究了溫和熱處理對腸炎沙門氏菌單細胞生長延滯期的影響,結果顯示隨熱處理溫度升高,單細胞生長延滯期的分布模型曲線范圍更廣,證明熱損傷增強了單細胞的生長變異性。此外,許多學者致力于非熱處理或其與熱處理相結合方式所致的食源性致病菌損傷及后續(xù)修復的研究。
Sibanda等采用流式細胞儀分選出經(jīng)酸(pH 4.2)、滲透壓(質量分數(shù)10%NaCl)及熱(55℃)處理后的單核細胞增生李斯特菌損傷群體并研究其生長延滯期,結果表明生長延滯期受菌株變異性(P<0.000 1)、處理條件(P=0.007)及修復溫度(P<0.001)的極顯著影響。此外,Kimura等應用400~600 MPa的高靜水壓處理大腸桿菌ATCC 25922并研究其損傷及修復動力學,發(fā)現(xiàn)在氧化應激強度更低的條件下生長延滯期更短,從而反映出經(jīng)高靜水壓處理后大量損傷菌的存在。McKellar等采用47℃熱處理與不同生長溫度(10~30℃)研究損傷程度及修復溫度對熒光假單胞菌生長延滯期及此過程中rRNA的rrnB P2啟動子活性的影響,結果顯示隨熱處理時間縮短及生長溫度的升高,生長延滯期縮短且rrnB P2啟動子的活性顯著提高,證明亞致死損傷會影響延滯期過程中基因的表達。Ma Jingjing等研究高壓處理損傷的Escherichia coliO157:H7修復動力學,結果也證明損傷菌會在生長延滯期過程中發(fā)生一系列生理及形態(tài)學變化并最終恢復至正常水平。下表總結了不同處理條件下食源性致病菌損傷與修復動力學的相關研究。
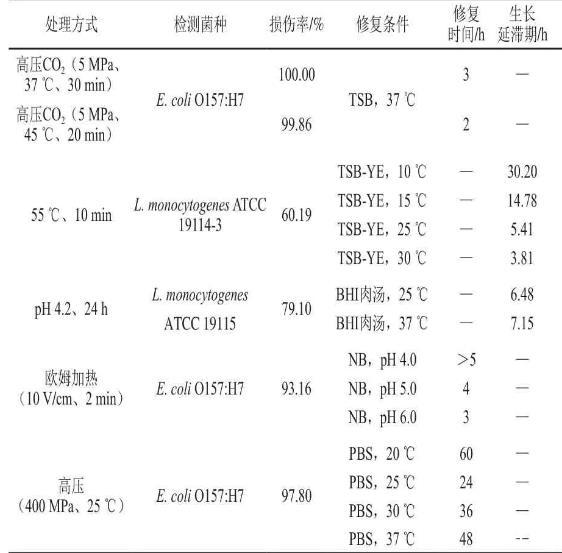
不同處理條件下食源性致病菌損傷與修復動力學的相關研究
亞致死損傷菌的存在會造成傳統(tǒng)致病菌檢測中污染水平的低估并且呈現(xiàn)更長的生長延滯期,在今后的研究中開發(fā)更準確、簡單、快速的方法來檢測損傷菌,并研究不同損傷及修復條件對生長延滯期修復過程的影響,對食品安全檢測及降低食品安全風險尤為重要。
初始接種細菌數(shù)量
傳統(tǒng)的預測微生物學研究認為初始接種細菌數(shù)量(>103CFU/mL)對食源性致病菌的生長沒有影響。然而,許多研究發(fā)現(xiàn)當食源性致病菌處于臨界狀態(tài)如接近生長邊界的溫度、pH值及滲透壓等條件或經(jīng)過歷史條件的脅迫壓力處理后,初始接種細菌數(shù)量會對生長延滯期造成一定的影響,尤其是在低接菌量條件下這種影響會更加顯著。Robinson等研究指出在含有1.2 mol/L NaCl溶液的TSB培養(yǎng)液中,單核細胞增生李斯特菌生長延滯期的均值及標準差均隨初始接種細菌數(shù)量的降低而增加,而未處理組單核細胞增生李斯特菌的生長延滯期不受接菌量的影響。Pin等通過隨機建模的方法探究不同初始接種細菌數(shù)量對大腸桿菌生長延滯期的影響,結果表明初始接種細菌數(shù)量越少則生長延滯期越長。類似結果也出現(xiàn)于Augustin等的研究。
食品中致病菌的污染大多是低數(shù)量級水平,并且其同樣具有適應食品中脅迫環(huán)境并生長繁殖的能力。因此,準確預測低菌量的致病菌在不同環(huán)境脅迫條件影響下的生長參數(shù)是必要的。董慶利等建立了單細胞生長流動成像系統(tǒng),并通過隨機建模的方法建立單細胞與群體細胞之間生長延滯期的關系,同時采用多次模擬探究不同初始接種細菌數(shù)量對銅綠假單胞菌群體細胞生長延滯期的影響,發(fā)現(xiàn)隨初始接種細菌數(shù)量的增加其生長延滯期縮短。類似的研究也見于Alonso等的報道,該研究通過建立隨機微分方程模型來描述單細胞生長與分裂的變異性,并通過模擬群體細胞生長,發(fā)現(xiàn)群體細胞生長延滯期隨初始接種細菌數(shù)量增加而縮短且變異性降低。
Aguirre等測定不同低接菌量下英諾克李斯特氏菌的生長延滯期,結果表明生長延滯期與初始接種細菌數(shù)量及生長溫度呈反比,與熱應激處理時間呈正比,且初始接種細菌數(shù)量對生長延滯期的影響既有隨機性因素又有微生物的生理因素。其中隨機性因素體現(xiàn)在單細胞生長延滯期的變異性以及影響該變異性的所有因素,如歷史條件造成細胞的亞致死損傷、接近生長邊界的實際生長條件等。另外,生理因素主要包括Kaprelyants等提出的“細胞間交流”,該研究表明原核生物細胞會使用某種(些)促生長的信號分子進行信息的傳遞,增加初始接種細菌數(shù)量會有利于這些信號分子的釋放與接收,從而縮短群體細胞的生長延滯期。
綜上所述,初始接種細菌數(shù)量與歷史、實際環(huán)境條件對食源性致病菌生長延滯期及變異性存在復雜的交互影響作用,研究以上交互效應如何影響生長延滯期對準確預測與控制食源性致病菌在食品中的生長至關重要,并且由于初始接種細菌數(shù)量顯著影響生長延滯期及變異性,在微生物定量風險評估的研究中需將初始污染水平考慮在內。
相關新聞推薦
1、呼寧散對雞肺源大腸桿菌生長曲線、細胞壁的影響及抑制效果——材料與方法