量化人類腸道菌種對艱難梭菌長期生長的影響(四)
關鍵的突變在艱難梭菌中顯著改變了代謝活動
因為進化型艱難梭菌的突變可能影響代謝活動,我們對高碳水化合物環境下進化型和原始型艱難梭菌菌株的exo-代謝組學特征進行了表征(見圖3B和S7F),以及限碳水化合物環境下的exo-代謝組學特征(見圖S5)。盡管原始菌株在代謝方面已知能夠利用脯氨酸,但進化型菌株在基礎培養基中并未顯著消耗脯氨酸(見圖3B)。相反,進化型菌株比原始菌株更有效地利用甘氨酸、天冬氨酸和N-乙酰甘氨酸。與此相反,進化型艱難梭菌在限碳水化合物培養基中的單菌種和與PV共培養條件下,脯氨酸的利用率顯著降低,其他氨基酸如甘氨酸、亮氨酸、異亮氨酸、纈氨酸和絲氨酸的消耗率顯著提高(見圖S5D)。此外,在兩種培養基條件下,進化型菌株的葡萄糖消耗量也高于原始菌株(見圖3B和S5D)。
為了探討這些突變如何影響艱難梭菌基因表達譜,我們在基礎培養基中無PV和有PV條件下,對原始型和進化型艱難梭菌菌株進行了基因組范圍轉錄組分析(見圖3C、S9A和S9B)。在3,508個總基因中,89個和55個在進化型艱難梭菌中顯著高表達和低表達,分別高于原始菌株(見圖3D和3E;表S8)。PTS相關基因,包括突變的基因206,在進化型菌株中顯著高表達。這表明,進化型菌株在轉運和磷酸化碳水化合物方面適應性更強,促進了如葡萄糖等碳水化合物的高效利用(見圖3B)。盡管prdR表達變化不大(1.4倍上調),但prd和grd操縱子的表達發生了顯著變化。特別是甘氨酸還原酶編碼基因顯著上調(2.1-至305倍),而脯氨酸還原酶編碼基因則顯著下調(3.0-至9.7倍),與原始菌株相比(見圖3E)。這一結果與exo-代謝組學結果一致,顯示了進化型菌株在脯氨酸和甘氨酸利用方面的變化(見圖3B)。
一些代謝基因,如負責Wood-Ljungdahl路徑(WLP)碳羰基分支的一氧化碳脫氫酶/乙酰-CoA合酶復合體,能夠將CO?轉化為乙酰-CoA,在進化型菌株中比原始菌株高表達(見圖3E)。WLP是最重要的能量產生和碳固定代謝之一,其中兩個CO?分子作為終端電子受體被還原為乙酰-CoA,進而轉化為乙酸。除此之外,包括thlA1、hbd、crt2(將乙酰-CoA轉化為丁酰-CoA)以及ptb和buk(將丁酰-CoA轉化為丁酸)的生產丁酸基因在進化型菌株中顯著高表達。與這些基因表達模式一致,進化型菌株丁酸生產能力比原始菌株高出2.7倍(見圖S9G)。作為一個參考,這種增強的丁酸生產能力(11.6 mM)與在類似培養基中培養的主要丁酸生產菌科普羅科科斯(Coprococcus comes)相當,可能在體內影響疾病嚴重程度。此外,先前的研究表明某些擬桿菌屬顯示出對丁酸的營養特異性生長敏感。在3種擬桿菌屬中(PV、BT、BU),只有PV在響應中等濃度的丁酸時顯示出顯著的生長抑制。進化型菌株產生的丁酸濃度位于PV丁酸-劑量響應曲線的低抑制區域(見圖S9H)。
在與PV共培養時,無論是原始菌株還是進化型艱難梭菌菌株均上調了丁酸發酵基因,而進化型菌株的這些基因表達更高(見圖S9C–S9E;表S9–S11)。由于培養基中不含丁酸,這表明了PV的交叉喂養現象,與有機酸測量結果的模式一致(見圖2C和2D)。值得注意的是,在進化型艱難梭菌菌株存在下,PV顯示出了許多與氨基酸生物合成相關的基因高表達,而與原始菌株相比(見圖S7B和S9F)。進化型艱難梭菌菌株消耗了比原始菌株更多的氨基酸(甘氨酸、纈氨酸、絲氨酸、亮氨酸和異亮氨酸)(見圖S5D)。這表明,PV向進化型艱難梭菌菌株的氨基酸交叉喂養可能比與原始菌株相比得到了增強(見圖S7B和S7C)。
進化型艱難梭菌菌株的生長受甘氨酸限制較小,對高葡萄糖濃度表現出增強反應
一個關鍵未解的問題是,這些代謝適應如何影響艱難梭菌在不同環境中的適應性(fitness)。為了評估適應性變化,我們研究了進化型和祖先型艱難梭菌菌株在不同氨基酸濃度下的生長情況。在氨基酸濃度降低(20%)的早期時間點,進化型艱難梭菌菌株的生長表現出顯著的優勢,而在其他條件下,兩者的生長反應類似(圖S7D)。
進化型艱難梭菌菌株的脯氨酸EC50(脯氨酸濃度,能夠達到最大生長的50%)顯著低于祖先型菌株(圖S10A和S10E)。這一結果表明,進化型菌株在低脯氨酸濃度下的競爭效率顯著高于祖先型菌株。脯氨酸在我們實驗條件下是祖先型艱難梭菌菌株的生長限制資源,這與其他具有多樣基因組的艱難梭菌菌株一致。然而,進化型菌株對脯氨酸的需求顯著低于祖先菌株,正如我們代謝組學數據(圖3B)所示。與這一結果一致,在PV共培養中,進化型菌株的豐度不隨脯氨酸濃度變化,這表明其生長處于脯氨酸濃度的飽和狀態(圖S10G)。相比之下,祖先型菌株的豐度隨著脯氨酸濃度的增加而增加(即脯氨酸劑量反應的線性狀態)。在低初始脯氨酸濃度下,進化型艱難梭菌菌株在12小時共培養中的豐度高于祖先型菌株,最終在實驗結束時下降(圖3F)。由于進化型菌株比祖先菌株更能消耗甘氨酸(圖3B),我們還研究了其對不同濃度甘氨酸的生長反應。在單培養(圖S10B和S10D)以及共培養PV(圖S10H)條件下,進化型和祖先菌株的生長響應顯示出類似的趨勢。
雖然進化型艱難梭菌菌株的葡萄糖EC50值高于祖先菌株,但進化型菌株在高葡萄糖濃度下表現出更高的生物量(即更高的最大生長產量,圖S10C和S10F)。這種生物量的增加并未通過單細胞水平上細胞大小和形態的變化來解釋(圖S7E)。這些發現暗示了在葡萄糖的生長產量和敏感性之間存在權衡。總之,進化型艱難梭菌菌株的代謝活動改變影響了其在不同脯氨酸和葡萄糖組合環境中的適應性(圖3G、3H和S11)。在高葡萄糖和低脯氨酸濃度條件下,相比祖先菌株,進化型菌株的適應性得到了增強。在某些環境中,這種適應性改變可能使進化型菌株在定殖過程中優于其他菌株,從而可能影響疾病結果。
進化型艱難梭菌菌株顯示出與人類腸道菌群的種間交互作用的變化
艱難梭菌代謝的顯著變化將影響人體腸道微生物群種間的交互作用。為了探討這些關鍵突變如何影響生態網絡,我們構建了包含進化型或原始艱難梭菌菌株的2-8成員群落的96種組合。其他種類(CS、CH、DP、BT、PV、BU和CA)在長期培養實驗中使用,廣泛分布于不同個體之間,涵蓋了人類腸道微生物群的系統發育多樣性,并在不同環境中被廣泛研究(圖4A)。我們將gLV模型擬合到物種絕對豐度的時間序列數據(表S3 DATASET002;圖S12A、S13A和S13B)。為了評估模型對未使用數據的預測性能,我們進行了20倍交叉驗證(圖S12B;STAR Methods)。模型預測與測得的物種豐度之間的吻合度良好(Pearson’s R=0.95–0.99,p<10E?05)。
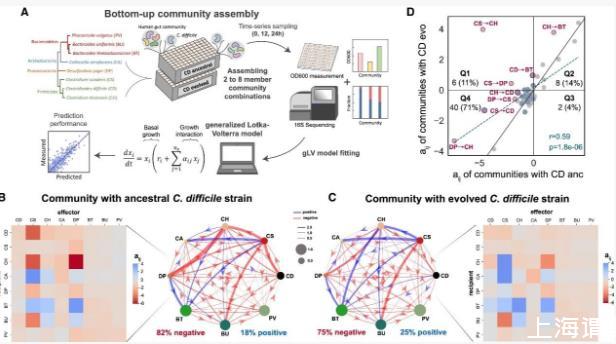
圖4|關鍵代謝基因突變改變了艱難梭菌與人體腸道微生物群種間的交互作用
(A)工作流程示意圖,用于解析艱難梭菌祖先株和進化株與人體腸道微生物之間的交互作用(STAR方法;表S3 DATASET002)。
(B和C)7種腸道微生物與祖先(B)和進化(C)艱難梭菌株之間的推斷種間交互網絡。節點大小代表各物種的載體容量,邊的寬度表示種間交互系數(aij)的大小。熱圖顯示了社區中8種物種之間的aij。
(D)祖先株與進化株艱難梭菌株的社區之間aij的散點圖。灰色(藍色)數據點是兩種腸道微生物之間(艱難梭菌與腸道微生物)的aij。藍色虛線表示兩社區aij值之間的線性回歸。兩側皮爾遜相關系數r和p值顯示。
相關新聞推薦
1、RIP 衍生物對?耐甲氧西林金黃色葡萄球菌生長曲線及生物膜形成的影響(一)
2、臨床結核分枝桿菌體外藥敏試驗:分子DST檢測VS表型DST藥敏診斷技術(一)