利用細菌生長曲線表征芳樟醇對三文魚莓實假單胞菌MS 02的抑菌效果(二)
2結果與分析
2.1α多樣性分析結果
對單樣本的α多樣性分析可以反映樣本內的微生物群落的豐富度和多樣性。由表1可知,5個樣品的微生物覆蓋率(Coverage)均為1.000,說明該測序結果鑒定了三文魚片中絕大多數細菌的系統類別。檢測結果表明,從0~12 d的樣品觀察到的細菌OTUs數量分別為121、104、125、135、128。以Shannon、Simpson、Chao1及Ace指數對樣品的菌群豐富度和多樣性進行初步評估。Shannon指數及Simpson指數用于估算樣品中微生物的多樣性,Shannon越大,群落多樣性越高,Simpson指數則相反;而Chao1指數及Ace指數常用于估算樣品的菌群豐度,是OTUs豐度的其他估計量。表中數據顯示Shannon指數逐日增加,而Simpson指數則相反,說明樣品的微生物多樣性逐漸增多;而Chao1及ACE指數均高于每個相應樣本的OTUs觀察數則表明在所有樣品中可能還存在一些額外的微生物門類。
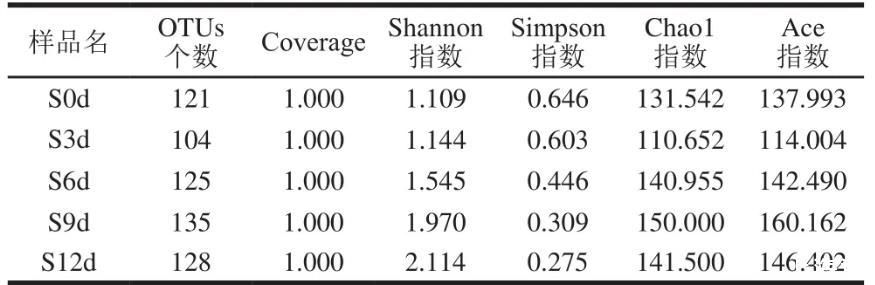
表1不同貯藏時間三文魚微生物群落的α多樣性指數
基于OTUs分析制作三文魚樣品的Venn圖,不同的橢圓代表不同組樣品,重疊部分的數字代表樣本間共有的OTUs個數,單獨的數字代表樣本特有OTUs個數。由圖1可知,整個貯藏期間三文魚OTUs總數為210個,其中5組樣品共有的有56個,占總數的26.67%;第0天(S0d)OTUs數量為121個,占總數的57.62%;第3天(S3d)OTUs數量為104個,占總數的49.52%;第6天(S6d)OTUs數量為125個,占總數的59.52%;第9天(S9d)OTUs數量為135個,占總數的64.29%;第12天(S9d)OTUs數量為128個,占總數的60.95%。第0天特有的OTUs數量為10個,占總數的4.76%;第3天特有的OTUs數量為4個,占總數的1.90%;第6天特有的OTUs數量為10個,占4.76%;第9天特有的OTUs數量為11個,占總數的5.24%;第12天特有的OTUs數量為12個,占總數的5.71%。圖中各橢圓均與其他組橢圓有重疊的部分,表明各組樣品的細菌物種之間互有交叉,圖1證明了在貯藏期間的三文魚細菌群落是動態變化的。
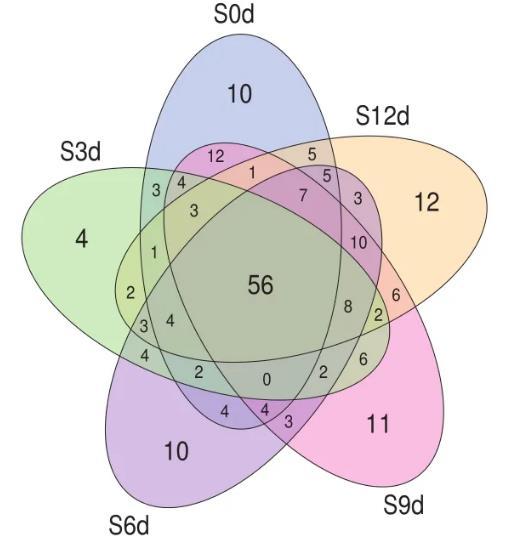
圖1三文魚樣品細菌群落Venn圖
2.2細菌相對豐度分析結果
分別從門、綱、目、科、屬、種6個水平對三文魚4℃貯藏期間的細菌群落相對豐度進行分析,結果如圖2所示。貯藏期間三文魚的細菌優勢菌門為變形菌門(Proteobacteria);優勢菌綱為γ-變形菌綱(Gammaproteobacteria);優勢菌目為弧菌目(Vibrionales)和假單胞菌目(Pseudomonadales)。在科水平的優勢菌為弧菌科(Vibrionaceae)、假單胞菌科(Pseudomonadaceae)、伯克氏菌科(Burkholderiaceae)和莫拉菌科(Moraxellaceae)。
三文魚4℃貯藏期間屬水平的優勢菌為發光桿菌屬(Photobacterium)、假單胞菌屬(Pseudomonas)、青枯菌屬(Ralstonia)和不動桿菌屬(Acinetobacter),相對豐度為97%,其中貯藏前期發光桿菌屬的細菌在整個菌相系統中處于優勢地位,相對豐度為84.89%;而隨著時間的延長,假單胞菌屬、青枯菌屬、不動桿菌屬的細菌相對豐度均呈現上升的趨勢,其中假單胞菌屬的細菌相對豐度上升趨勢最為明顯,在貯藏初期相對豐度為3.98%,到第12天時相對豐度高達31.46%,并且仍有上升的趨勢。此外菌相系統中還存在一些如紫色桿菌屬(Janthinobacterium)、希瓦氏菌屬(Shewanella)等細菌。而在整個貯藏期間Pseudomonas fragi增長速度最快,在第0、3、6、9、12天相對豐度分別為3.01%、4.89%、11.20%、21.99%、24.82%。
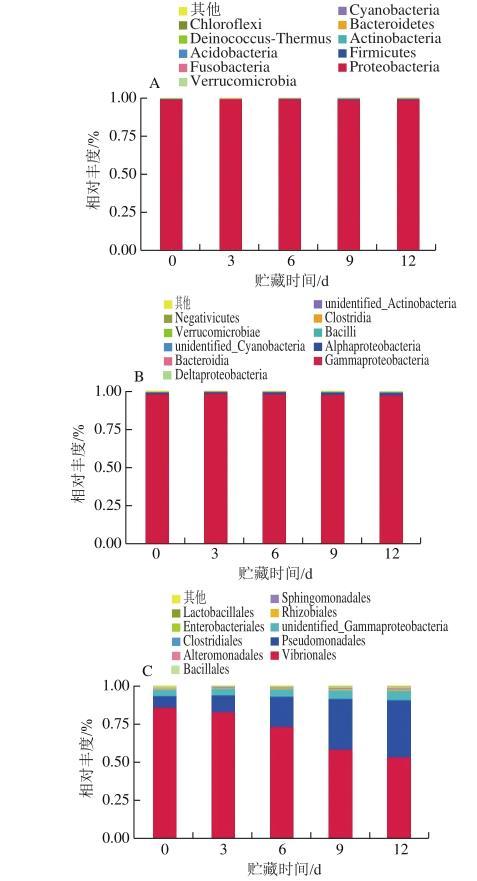
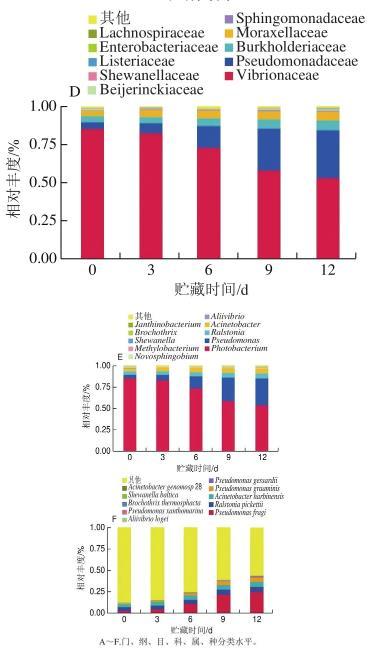
圖2三文魚細菌群落不同分類水平的相對豐度
2.3優勢腐敗菌的分離鑒定結果
將1.3.2節分離、純化后得到菌株進行DNA提取和PCR擴增,測定擴增產物的基因序列并與NCBI數據庫中已知的微生物基因序列進行BLAST比對分析,構建系統發育樹,選擇與已知的微生物基因序列同源性最高且活性最強(活化培養至對數生長期時菌體濃度最高)的菌株,將其命名為MS 02。如圖3所示,16S rRNA分析結果證明MS 02為莓實假單胞菌。后續實驗均以莓實假單胞菌MS 02為靶細菌。

圖3菌株MS 02與其他假單胞菌之間的系統發育樹
2.4芳樟醇對莓實假單胞菌的抑菌活性
芳樟醇對水產品優勢腐敗菌——莓實假單胞菌的抑菌活性如表2所示。當芳樟醇質量濃度為1.00μg/mL時,莓實假單胞菌的OD595 nm為0.268±0.014,與陽性對照組無顯著性差異(P>0.05),但略高于陽性對照組,因此芳樟醇對莓實假單胞菌的MIC為2.00μg/mL。圖4顯示了芳樟醇對莓實假單胞菌MS 02生長的影響,在細菌生長過程中,菌懸液中的細菌濃度與OD595 nm呈正相關,因此能夠通過莓實假單胞菌OD595 nm表征細菌的生長狀況。由圖4可知,空白對照組莓實假單胞菌呈指數型增長,說明溶劑中的甲醇對細菌生長無明顯影響。加入MIC和2 MIC的芳樟醇后,細菌生長明顯受到抑制,MIC組的OD595 nm僅在培養后期有略微增長,可能是因為芳樟醇劑量較低,且培養時其易揮發導致的。
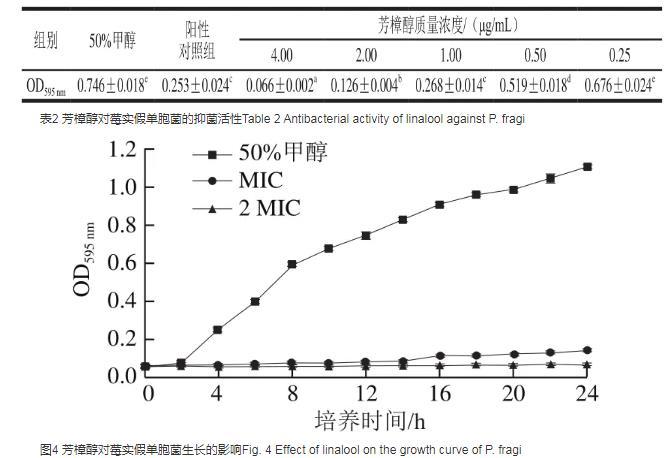
圖4芳樟醇對莓實假單胞菌生長的影響
2.5莓實假單胞菌的微觀形貌觀察結果
采用芳樟醇處理后的莓實假單胞菌的微觀形貌如圖5所示,未添加芳樟醇的莓實假單胞菌(圖5A)的菌體形態呈完整且表面光滑的桿狀結構,當添加芳樟醇后,菌體表面隨著芳樟醇添加量的增大出現不同程度的破裂,MIC和2 MIC芳樟醇處理的莓實假單胞菌如圖5B、C所示,圖5B中菌體表面變得粗糙且不完整,細胞膜產生明顯的凹陷褶皺,圖5C中的菌體形態被嚴重破壞,細胞膜幾乎完全破裂,內容物溶出。上述結果表明芳樟醇能破壞莓實假單胞菌的細胞結構,導致細胞內容物外泄,從而抑制菌的生長。植物精油因具有疏水性更易作用于細菌細胞膜,破壞胞質膜結構,從而抑制細菌生長。
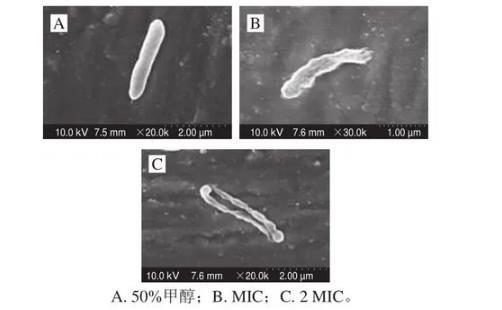
圖5芳樟醇處理后的莓實假單胞菌SEM圖
2.6芳樟醇對莓實假單胞菌電導率的影響
圖6反映了芳樟醇對莓實假單胞菌MS 02電導率的影響,當細菌細胞膜被抑菌劑損害時,細胞膜的半透性喪失,胞內電解質大量流出,抑制細菌生長。空白對照組菌液電導率變化不明顯,這表明體系中的甲醇不會破壞細胞膜的通透性;加入MIC和2 MIC芳樟醇的菌液電導率迅速上升,表明芳樟醇對莓實假單胞菌的細胞膜有明顯破壞作用,細胞膜破損、電解質流出導致菌液電導率的上升。
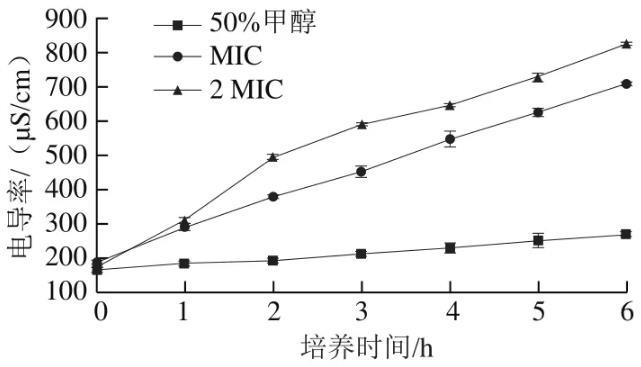
圖6芳樟醇對莓實假單胞菌電導率的影響
2.7芳樟醇對莓實假單胞菌細胞膜通透性的影響
FDA是一種自身無熒光且不帶電荷的脂質性分子,在胞內可被酶水解為熒光素發出熒光,當細胞膜受到破壞,熒光素流失從而導致FDA熒光強度降低。因此,細菌細胞膜的通透性可通過FDA熒光強度來反映。圖7中顯示的是相同處理時間不同處理條件的FDA熒光強度,50%甲醇處理的熒光強度為1 920 AU,MIC和2 MIC組的熒光強度分別為530、211 AU,可以看出添加芳樟醇的處理組其熒光強度明顯低于對照組,這表明50%甲醇不能改變細菌細胞膜的通透性。但隨著芳樟醇質量濃度增加,細菌細胞膜被破壞程度逐漸明顯,2 MIC處理組的熒光強度最低,這表明2 MIC芳樟醇處理能明顯破壞莓實假單胞菌的細胞膜,這與圖5中2 MIC組菌體形態被嚴重破壞,細胞膜幾乎完全破裂的結論一致。舒慧珍等在探究檸檬烯對熒光假單胞菌細胞膜通透性的影響時也得到了類似的結論。
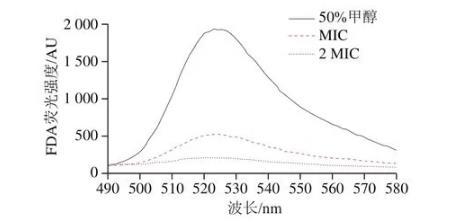
圖7芳樟醇對莓實假單胞菌FDA熒光強度的影響
2.7芳樟醇對莓實假單胞菌細胞膜完整性的影響
細胞膜的完整性是菌體正常生長代謝的重要影響因素,其受到破壞會導致膜內的DNA和RNA等物質泄漏,DNA和RNA在260 nm波長處具有強吸收作用,故可用菌懸液在OD260 nm的變化反映細菌細胞膜的完整性。如圖8所示,在反應3 h后,含芳樟醇組菌懸液中的OD260 nm迅速升高,而空白對照組變化不明顯,這與上文的結論相一致,說明PCL不會引起細菌細胞膜的破裂。MIC和2 MIC的芳樟醇會使莓實假單胞菌細胞膜在短時間內破裂,內容物迅速流失,對細菌細胞有較明顯的破壞作用。張赟彬等發現肉桂醛會使大腸桿菌和金黃色葡萄球菌的細胞膜完整性被破壞,內容物迅速釋放導致菌液在260 nm波長處的光密度值增大,本實驗結果與其一致。
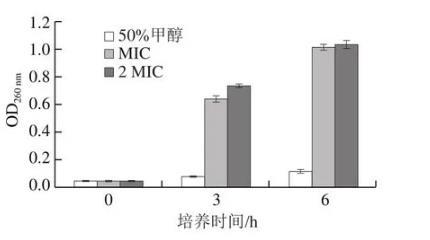
圖8芳樟醇對莓實假單胞菌細胞膜完整性的影響
2.8芳樟醇對莓實假單胞菌胞內酶的影響
鈉鉀ATP酶是一種蛋白酶,存在于組織細胞及細胞器膜上,能維持跨膜離子濃度梯度,測定胞內鈉鉀ATP酶的活力變化可以分析抑菌劑對細菌細胞能量代謝的影響。由圖9可知,在培養6 h后,空白對照組的鈉鉀ATP酶活力最大,而MIC和2 MIC芳樟醇組的鈉鉀ATP酶活力都相對較小,且明顯可以看出芳樟醇添加量與胞內鈉鉀ATP酶活力有直接聯系。這表明芳樟醇能有效抑制莓實假單胞菌細胞內鈉鉀ATP酶的活力,影響細胞的正常能量代謝。胡文杰等研究表明油樟葉精油的幾種餾分單體也能使尖孢鐮刀菌內ATP酶活力下降。
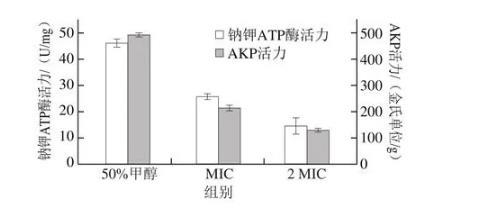
圖9芳樟醇對莓實假單胞菌胞內酶活力的影響
AKP存在于細胞壁和細胞膜之間,一般難以在胞外被檢測到,但當細胞結構被破壞后,AKP會泄漏至細胞外。因此,可通過檢測細菌胞外的AKP活力變化來觀察其對細胞壁的破壞情況。由圖9可知,在培養6 h后,空白對照組AKP活力最高,說明50%甲醇對莓實假單胞菌的細胞壁結構無影響。但添加MIC和2 MIC芳樟醇后,莓實假單胞菌AKP活力降低,這表明芳樟醇的加入會使莓實假單胞菌的細胞壁被破壞,導致菌體電解質外泄,最終抑制菌體生長,且隨著芳樟醇添加量的增加,AKP活力降低越明顯。
3結論
本實驗采用HTS分析4℃貯藏期間三文魚的菌相結構,并分離提取出1株三文魚優勢腐敗菌——莓實假單胞菌MS 02,再深入探究芳樟醇對莓實假單胞菌MS 02的抑菌機理。4℃貯藏三文魚的微生物群落多樣性與演替規律會隨著時間延長而變化。三文魚的優勢菌屬為發光桿菌屬、假單胞菌屬、青枯菌屬和不動桿菌屬。貯藏期間三文魚菌相發生極大地變化,各菌群之間相互競爭,假單胞菌屬的相對豐度隨著時間的延長逐漸增大,莓實假單胞菌是樣品在貯藏期間的增長速率最快的菌種。芳樟醇對莓實假單胞菌MS 02具有良好的抑菌效果,SEM結果表明芳樟醇能使細菌細胞膜產生凹陷褶皺,而電導率、OD260 nm、FDA熒光強度和AKP、鈉鉀ATP酶活力結果則證明了芳樟醇能破壞莓實假單胞菌的細胞膜和細胞壁的通透性和完整性,影響細菌細胞正常能量代謝,導致菌體死亡。
利用細菌生長曲線表征芳樟醇對三文魚莓實假單胞菌MS 02的抑菌效果(一)
利用細菌生長曲線表征芳樟醇對三文魚莓實假單胞菌MS 02的抑菌效果(二)